The ISME Journal (2 0 1 4 ), 1 -1 2
© 2 0 1 4 International Society for Microbial Ecology All rights reserved 1 7 5 1-7 362 /14
OPEN
ww w .nature.com /ism ej
ORIGINAL ARTICLE
Natural occurrence of microbial sulphur oxidation
by long-range electron transport in the seafloor
Sairah Y M alkin1,2,4, A lexandra MF Rao1,5, Dorina Seitaj2, Diana Vasquez-Cardenas2,3,
Eva-Maria Zetsche1, Silvia Hidalgo-M artinez2, Henricus TS Boschker
and Filip JR M eysm an1,2
1Department o f Analytical, Environm ental, and Geo-chemistry, Vrije Universiteit Brussel (VUB), Brussels,
Belgium; 2D epartment o f Ecosystem Studies, Boyal Netherlands Institute fo r Sea Besearch (NIOZ), Yerseke,
The Netherlands and 3Department o f Marine Microbiology, Boyal Netherlands Institute fo r Sea Besearch
(NIOZ), Yerseke, The Netherlands
Recently, a novel mode of sulphur oxidation was described in marine sediments, in which sulphide
oxidation in deeper anoxic layers was electrically coupled to oxygen reduction at the sediment
surface. Subsequent experimental evidence identified that long filamentous bacteria belonging to
the family Desulfobulbaceae likely mediated the electron transport across the centimetre-scale
distances. Such long-range electron transfer challenges some long-held views in microbial ecology
and could have profound implications for sulphur cycling in marine sediments. But, so far, this
process of electrogenic sulphur oxidation has been documented only in laboratory experiments and
so its imprint on the seafloor remains unknown. Here we show that the geochemical signature of
electrogenic sulphur oxidation occurs in a variety of coastal sediment environments, including a salt
marsh, a seasonally hypoxic basin, and a subtidal coastal mud plain. In all cases, electrogenic
sulphur oxidation was detected together with an abundance of Desulfobulbaceae filaments.
Complementary laboratory experiments in intertidal sands demonstrated that mechanical dis­
turbance by bioturbating fauna destroys the electrogenic sulphur oxidation signal. A survey of
published geochemical data and 16S rRNA gene sequences identified that electrogenic sulphide
oxidation is likely present in a variety of marine sediments with high sulphide generation and
restricted bioturbation, such as mangrove swamps, aquaculture areas, seasonally hypoxic basins,
cold sulphide seeps and possibly hydrothermal vent environments. This study shows for the first
time that electrogenic sulphur oxidation occurs in a wide range of marine sediments and that
bioturbation may exert a dominant control on its natural distribution.
The ISME Journal advance online publication, 27 March 2014; doi:10.1038/ismej.2014.41
Subject Category: Geomicrobiology and microbial contributions to geochemical cycles
Keywords: bioturbation; Desulfobulbaceae; electrobiogeochemistry; electrogenic sulphur oxidation;
sulphur oxidizing bacteria
Introduction
Sulphate reduction is the dom inant m ineralization
pathw ay in coastal m arine sedim ents, and leads to
an accum ulation of sulphide in deeper sedim ent
horizons (Jorgensen, 1982). Various respiratory
pathw ays of m icrobial sulphur oxidation have
evolved as a m eans of harvesting this stored
Correspondence: SY M alkin or FJR Meysman, Analytical,
Environm ental, and Geo-chemistry, Vrije U niversiteit Brussel,
Pleinlaan 2, Brussels 1050, Belgium.
E-mail: smalkin@uga.edu or filip.meysman@nioz.nl
4Current address: Departm ent of Marine Sciences, U niversity of
Georgia, Athens, GA, USA.
5Current address: Institut des Sciences de la Mer (ISMER),
U niversité du Québec à Rimouski (UQAR), Rimouski, QC,
Canada.
Received 14 November 2013; revised 4 February 2014; accepted
16 February 2014
chem ical energy (Jorgensen and Nelson, 2004).
Recently, a novel sulphur oxidation process, herein
called electrogenic sulphur oxidation, was discov­
ered whereby the oxidation of free sulphide in
deeper sedim ent horizons was observed to be
directly coupled to reduction of oxygen near the
sedim ent surface (Nielsen et ah, 2010). Such a
spatial separation of redox half-reactions indicates
there m ust be a direct pathw ay by w hich electrons
originating from sulphide are conducted across
centim etre-scale distances to oxygen (Nielsen et al,
2010; Risgaard-Petersen et al., 2012). Experim ental
m anipulation revealed that this electrical connec­
tion responded im m ediately (that is, w ithin
m inutes) to oxygen depletion in the overlying
water, and so the connection was too fast to be
explained by diffusion of redox-active shuttles
(Nielsen et ah, 2010). M etal-reducing bacteria are
Long-range electron transport In th e seafloor
SY M alkin e f a/
know n to transport electrons externally via cell
surface cytochromes, redox shuttles or conductive
pili (Reguera et al., 2005; Gorby et al., 2006; Lovley,
2008; Clarke et al., 2011), and such extracellular
electron transport can bridge ~ 100 m icrometer
distances in electrode biofilms (Reguera et al.,
2006; Logan and Rabaey, 2012). The electron trans­
port associated w ith this electrogenic sulphur
oxidation, by contrast, operates on scales of centi­
metres, thereby extending the known length scale of
microbially m ediated electron transport by two
orders of m agnitude.
Electrogenic sulphur oxidation creates a unique
geochemical signal in surface sedim ents, w hich is
detectable by m icrosensor profiling. A key feature is
that the cathodic half-reaction (that is, 0 2 + 4e~ +
4H + ->2H20 ) consumes protons, thereby creating a
pH m axim um in the oxic zone. Deeper in the
sedim ent, protons are released by an anodic half­
reaction (that is, electrogenic sulphide oxidation;
such as V2 H2S + 2H20->V2S0t~ + 4 e ~ + 5 H + ) creat­
ing a broad pH m inim um near the sulphide horizon,
separated from the oxic zone by a suboxic zone
ranging from several m illim etres to approxim ately
2 cm wide. In laboratory experim ents, the develop­
m ent of this characteristic geochemical fingerprint
has been used as a sign of electrogenic sulphur
oxidation activity (Nielsen et al., 2010; Pfeffer et al.,
2012; Risgaard-Petersen et al., 2012).
Compelling evidence indicates that electrogenic
sulphur oxidation is likely carried out by long
filam entous bacteria. First, in sedim ents w ith the
geochemical signature of this process, detailed
m icroscopic investigation revealed high densities
of long m ulticellular filam entous bacteria, belonging
to the family Desulfobulbaceae, w hich were span­
ning the entire length of the suboxic zone (Pfeffer
et al., 2012). Second, w hen a thin wire was passed
horizontally through the suboxic zone, the electric
circuitry was effectively ‘cut’, as indicated by an
im m ediate decrease in sedim ent oxygen consum p­
tion by a factor of 2, and a decrease in the signature
pH maximum. This experim ent identified that the
electron conduction occurred along a continuous
structure spanning the suboxic zone (Pfeffer et al.,
2012). Thirdly, w hen sedim ent was experim entally
incubated w ith filters of various pore sizes w hich
were inserted horizontally, the geochemical signa­
ture of electrogenic sulphur oxidation developed
only in cores that had filters w ith pore sizes large
enough for Desulfobulbaceae filaments to penetrate
(Pfeffer et al., 2012). Taken together, these experi­
ments provide strong evidence that electrogenic
sulphur oxidation is carried out by Desulfobulbaceae
filaments.
The capacity of Desulfobulbaceae filaments
to oxidize su lp h u r by long-range electron
transport represents an entirely novel m icrobial
lifestyle (Pfeffer et al., 2012). By conducting
electrons across centim etre-scale distances, these
filam entous bacteria appear capable of enabling a
The ISME Journal
previously unrecognized strategy for resource
competition in marine sediments (Nielsen et al., 2010;
Risgaard-Petersen et al., 2012). Yet until now,
electrogenic sulphur oxidation has only been
induced in laboratory experim ents that have used
sieved and hom ogenized sedim ent from a deep
sedim ent horizon obtained from one particular
location in the Baltic Sea (Nielsen et al., 2010;
Pfeffer et al., 2012; Risgaard-Petersen et al., 2012).
Our objective here is to identify w hether
electrogenic su lp h u r oxidation, together w ith
D esulfobulbaceae filam ents, are found in natural
m arine sedim ents. We further aim to identify
major environm ental constraints on the occurrence
and distribution of electrogenic sulphur oxidation
in m arine sedim ents. Given the high growth rate and
densities of these bacteria observed in laboratory
experim ents (Pfeffer et al., 2012; Risgaard-Petersen
et al., 2012), we hypothesize that electrogenic
sulphur oxidation is a com petitively successful
strategy in natural m arine sedim ents and therefore
may be w idespread in nature.
Materials and methods
Site descriptions
The laboratory experim ents of Nielsen et al. (2010)
and Pfeffer et al. (2012) suggested that electrogenic
sulphur oxidation may be com petitively successful
in coastal m arine sedim ents w ith high rates of
sulphide generation linked to high rates of organic
m atter m ineralization. Our site selection therefore
targeted four different coastal seafloor habitats in the
southern North Sea area, three of w hich were in
depositional areas and were particularly rich in
organic matter. We exam ined intact sedim ents
collected from these sites for geochemical evidence
of electrogenic sulphur oxidation and for the
presence of Desulfobulbaceae filaments. Our first
site was Rattekaai Salt M arsh (RSM), an intertidal
area heavily loaded w ith organic matter, including
large periodic deposits of macroalgal detritus, w hich
supports high rates of sulphate reduction (Oenema,
1988; Pallud and Van Cappellen, 2006; Table 1).
We sam pled the un-vegetated sedim ents in an
intertidal drainage channel. At low tide, sulphiderich porewater seeps out from these creek sediments.
Large bioturbating anim als were absent from these
seepage areas, likely excluded by high levels of
porew ater sulphide. Our second site was Station 130
in the Belgian Coastal Zone (BCZ), a subtidal site at
12 m water depth (Table 1). This site is characterized
by high accum ulation rates of m uddy deposits
w hich generate intense sulphate reduction and free
sulphide was previously detected below a suboxic
zone of several centim etres (Gao et al., 2009).
A m onthly survey in 2011 revealed that large
bioturbating m acrofauna is generally absent from
this site (Braeckman et al., 2014), and no m acro­
fauna was detected in our collected sedim ent cores.
Long-range electron transport In the seafloor
SY M alkin e ta /
3
Table 1 The geochemical fingerprint of electrogenic sulphur oxidation was observed at three sites in the coastal North Sea under field
conditions
Site nam e
RSM, Oosterschelde, The
N etherlands
Station 130, BCZ
Coordinates
N
E
N
E
MLG, Holpervoet and Den Osse N
basins, The Netherlands
E
OSF, The Netherlands
Dates o f
sam pling
5T 26'
04° 10'
5T 16'
02° 54'
51“ 46'
03c 56'
21"
22 June 2011
11" 25 February 2013
13" 10 October 2011
19"
1 M arch 2013
24" 3 November 2011
16" 22 January 2012
24 February 2012
N 5T 26' 52"
17 May 2011
E 04° 05' 47" 15 August 2011
OPD
(mm)
0.7 ±0.1
1.3 10.4
0 .710.1
1.2 10.8
0 .9 1 0 .2
1 .4 1 0 .4
1.8 10.4
1.610.3
1.5 10.3
DOU
Current density
0 2 consum ed by
(m m ol m ~ 2 d ^ 1)
cathodic reaction (%)
(m A m ~ 2)
30 .3 1 3 .6
19.912.2
32.3115.0
24.8120.9
44.1116.1
13.914.1
22.2 19.5
4 .8 1 1 .6
3 .5 1 0 .5
2 9 .9110.7
7.3 14.0
25.2 12.6
4 .6 1 1 .8
24.8 15.5
16.1 110.4
27.0116.2
22.7110.1
8.2 1 4 .3
19.11 7.1
5 .3 1 2 .9
14.01 7.9
29.5125.3
33.71 30.6
NA
NA
Abbreviations: BCZ, Belgian Coastal Zone; DOU, diffusive oxygen uptake across the sediment-water interface; MLG, Marine Lake Grevelingen;
OPD, Oxygen Penetration Depth; OSF, Oosterschelde sand flats; RSM, Rattekaai Salt Marsh.
Values indicate means of between 2 and 4 replicate cores. Reported error is standard deviation between replicate cores.
Our third site was M arine Lake Grevelingen (MLG),
a seasonally stratified coastal m arine system in the
Delta area of The N etherlands w hich experiences
bottom water hypoxia and anoxia in the summ er in
its deep basins (Table 1). We collected sedim ents
from the deepest area of two steep-sided basins
(Holpervoet basin at 23 m; Den Osse basin at 35 m
water depth). These basins rapidly accum ulate
organic rich deposits ( > 2 cm per year) due to
focussing of detrital material. At the time of
sam pling (winter), the water colum n was fully
oxygenated, but even so, m acrofauna is never
abundant in these seasonally anoxic sedim ents and
no m acrofauna was present in our retrieved sedi­
m ent cores. The fourth site was an intertidal sand
flat in the Oosterschelde tidal inlet (Delta area, The
Netherlands). Unlike the first three sites, the
Oosterschelde Sand Flats (OSF) support high rates
of bioturbation, most notably by the lugworm
Arenicola marina (Coosen et al., 1994). Free
sulphide concentrations are low to non-detectable
(< 2 0 p m o ll_1) in at least the top 2.5 cm of the
sedim ent. All sites were visited on two or three
occasions (Table 1).
The intertidal sedim ent samples (that is, RSM and
OSF) were collected by m anually inserting core
tubes (4 cm diameter) in the sedim ent at low tide.
The sedim ent from MLG was retrieved w ith a
Uwitec gravity corer (6 cm diameter). On the first
occasion, sedim ent from the BCZ was collected w ith
a Reineck box corer followed by careful subsam pling
w ith a 10 cm diam eter core tube. On the second
occasion, sedim ent from this site was collected w ith
a Uwitec gravity corer. Retrieved cores were closely
inspected and only cores w ith an undisturbed
sedim ent surface were kept for the study.
M icroprofiling was either conducted im m ediately
on-board the sailing vessel (BCZ and MLG in 2011
only), or sedim ent cores were returned to a nearby
laboratory where m icroprofiling was conducted
under
clim ate-controlled
conditions.
Microprofiling was conducted betw een 2 and 12 h after
core collection under darkened conditions. The
intertidal sedim ents from RSM and OSF were kept
in the dark for at least 6 h prior to m icroprofiling to
ensure there was no effect of photosynthesis on the
pH profiles.
Microelectrode profiling and reactive transport
modelling
Sedim ent profiling was performed using commer­
cial m icrosensors operated w ith a m otorized m icro­
m anipulator (Unisense A.S., Aarhus, Denmark).
Sedim ent was brought level to the core liner surface.
Cores were placed in an aquarium containing water
collected from the study site and held at in situ
tem perature and constantly bubbled w ith air. During
m icrosensor profiling, an airstream was additionally
provided at the water surface upstream of the
sedim ent cores to ensure constant water flow over
the sedim ent, estim ated at 2 - 4 c m s -1 at 0.5cm
above the sedim ent surface. Oxygen and H2S
m icroelectrodes had a tip diam eter of 50 pm, and
pH m icrosensors had a tip diam eter of 100 or
200 pm. Vertical oxygen m icroprofiles were recorded
at 100 pm steps, beginning at least 1m m above the
sedim ent-w ater interface. A tw o-point 0 2 calibra­
tion was m ade in air-saturated seawater and the
anoxic zone of the sedim ent (100% and 0%,
respectively). Vertical H2S and pH m icroprofiles
were m ade concurrently, recorded at 200 pm steps in
the oxic zone, and 200 or 400 pm steps below. For
H2S, a five-point standard curve was m ade using
Na2S standards. ZH2S was calculated from H2S
based on pH m easured at the same depth using the
R package AquaEnv (Hofmann et al., 2010). For pH,
a three-point calibration using NBS standard buffers
was made, followed by a salt correction w ith TRIS
buffer (Dickson et al., 2007). pH m easurem ents were
perform ed w ith a Ag/AgCl reference electrode and
values are reported on the total scale. Porosity
was determ ined from water content and dry density
m easurem ents upon drying to constant weight
at 60 °C.
The ISME Journal
Long-range electron transport In th e seafloor
SY M alkin e f a/
Reactive transport m odelling was used to
calculate reaction rates from the m icrosensor data
(Table 1). The diffusive oxygen uptake (DOU) of the
sedim ent was calculated from the oxygen depth
profile near the sedim ent-w ater interface using
Fick’s first law as below:
Ji=q>/{ 1 - 2lnç>)Df(S, T)dC i/dx
where q> is the m easured porosity, the term (1 —2ln
<p) is a correction for sedim ent tortuosity (Boudreau,
1996), and D¡ is the diffusion coefficient for oxygen
calculated at salinity (S) and tem perature (T)
calculated using the R package marelac (Soetaert
et al., 2012). The m axim um concentration gradient,
dC¡/dx, was calculated as the linear slope im m edi­
ately below the sedim ent-w ater interface.
The cathodic proton consum ption (CPC) is the
proton consum ption corresponding to the cathodic
half-reaction, 0 2 + 4e~ + 4H + ->2H20 . The CPC was
calculated as the sum of the upw ard and dow nw ard
alkalinity fluxes im m ediately above and below the
pH m axim um in the oxic zone. A lkalinity fluxes
were calculated as the sum of the fluxes of the major
individual porew ater com pounds that contribute to
the alkalinity:
7 t A = / h C 0 3-
+ 2 /co2- + 7 b (OH)“
+ /
h s~
+ /
oh
7h +
The spéciation of individual ions (carbonate,
borate, sulphide and water equilibria) were calcu­
lated from H2S and pH m icrosensor data using the R
package AquaEnv (Hoffman et al., 2010). For this
calculation, we assum ed a constant depth profile
for dissolved inorganic carbon and borate. The
contribution of other m acronutrients (ammonium,
silicate) to alkalinity fluxes are likely small and
were not considered in this study.
The concentration gradient dQ/dx for each individual
species was estim ated by a noise robust num erical
differentiation using the Savitsky-Golay algorithm,
optim ized w ith a iV=21 point stencil (Savitsky and
Golay, 1964). The current density was calculated
from the CPC using a conversion factor of one
electron per proton (1A = 1.036 x 10~5m ole~ s _1).
The percentage, p, of oxygen consum ed electrochem ically was calculated as P = 25 (CPC/DOU),
based on a reaction stoichiom etry that four moles of
H + are consum ed per mole of 0 2 by the cathodic
reaction. The rem aining part of the oxygen
consum ption (1 —p) is attributed to aerobic respira­
tion, w hich does not consum e or release alkalinity.
This sim plified m odel assumes that no other
biogeochemical reactions are releasing or consuming
alkalinity w ithin the oxic zone. In reality, some
alkalinity consum ption will take place, associated
w ith nitrification, ferrous iron oxidation, calcium
carbonate precipitation or direct sulphur oxidation
by oxygen (Risgaard-Petersen et al., 2012). The
rates of these reactions cannot be estim ated from
m icrosensor profiles. However, correcting for
these reactions w ould increase the value of p.
Accordingly, the p estimates derived here should
The ISME Journal
be interpreted as conservative, lower bounds on the
oxygen that is consum ed electrochemically.
Identification of Desulfobulbaceae filaments
M icroscopic identification of Desulfobulbaceae
filaments was achieved by catalysed reporter
deposition-fluorescence in situ hybridization
(CARD-FISH; Pernthaler et al., 2002), using a
Desulfobulbaceae-specific oligonucleotide probe
(DSB706; 5'-ACC CGT ATT CCT CCC GAT-3')
labelled w ith horseradish peroxidase (Liicker et al.,
2007). Sedim ent samples were fixed w ith a 1:1
(vol/vol) ethanol solution and stored at —20 °C until
analysis. H undred m icrolitres of the preserved
sample sedim ent slurry was pipetted onto polycar­
bonate m em brane filters (0.2 pm pore size). The
retained filam entous cells were perm eabilized w ith
lysozyme ( l h at 37 °C) and treated w ith achromopeptidase (30m in at 37 °C). Endogenous peroxidases
were inactivated w ith a m ethanol and peroxide
solution (30m in at 20 °C), and subsequently hybri­
dized in standard hybridization buffer (46 °C) w ith
45% formamide. CARD-FISH was also performed
w ith probes EUB388-I and NONEUB388 (with 10%
formamide) as positive and negative controls,
respectively. Subsequently, hybridized cells were
visualized using epifluorescence
microscopy
(Axioplan II Imaging, Zeiss, Jena, Germany).
Desulfobulbaceae filaments were additionally quan­
tified from suboxic zone (0.3-0.8 cm) sedim ent
sections obtained from RSM, BCZ and OSF, using
the FISH probe as above. Filam ent length was
m easured in 200 random m icroscopic fields.
Sedim ent samples from MLG enabled filament
identification, but could not be used for enumeration.
Specim ens for scanning electron microscopy
(SEM) were prepared by isolating filaments w ith a
hook m ade from flame-drawn pasteur pipettes
under a light microscope ( x 40) and transferring
these filaments to polycarbonate filters (pore size
5 pm). Filam ents retained on filters were then
transferred through a gradual ethanol gradient
(25-100% ), critical point dried w ith liquid C 0 2,
m ounted on stubs and coated w ith gold. Images
were m ade w ith a JEOL 5600 SEM under lowvacuum, operated at 30 kV.
Partial 16S rRNA gene sequences of the
Desulfobulbaceae filaments were recovered by
constructing 16S rRNA cDNA clone libraries using
suboxic zone sections of sedim ents (0.3-0.8 cm) in
w hich the geochemical fingerprint of electrogenic
sulphide oxidation was detected. RNA was extracted
w ith a ZR Fungal/Bacterial RNA M iniprep kit
(Zymo Research, Irvine, CA, USA) and reversetranscribed using random prim ers. The 16S rRNA
gene was am plified by PCR using general bacterial
prim ers (Bac F968.60 and Bac R140.10), cloned into
Escherichia coli by one shot chem ical transform a­
tion and sequenced using a genetic analyzer
(3130 Genetic Analyzer, A pplied Biosystems, Grand
Long-range electron transport In the seafloor
SY M alkin e ta /
Island, NY, USA). Sequences were aligned w ith the
software ARB (Ludwig et al., 2004) against the
SILVA database. A phylogenetic tree was con­
structed in ARB by m axim um parsimony, w ith
1000 bootstrap iterations. The software Pintail was
used to check for chimeras (Ashelford et al., 2005).
Related 16S rRNA gene sequences were obtained by
BLAST searches of GenBank.
Experiments with bioturbated sediments
In addition to the field study, we perform ed two
laboratory experim ents to investigate the role of
bioturbation on the natural distribution of electro­
genic sulphur oxidation in coastal m arine sedi­
m ents. Sedim ents collected from the heavily
bioturbated OSF field site did not show evidence
of electrogenic sulphur oxidation or a detectable
presence of Desulfobulbaceae filaments on repeated
visits (details in Results). In a first experim ent, we
tested w hether the geochemical signature of electro­
genic sulphur oxidation could develop in these
sediments when bioturbation is removed. Sediments
from the sand flats differed from the other
three study sites in several ways, including a coarser
grain size and lower water content, lower organic C
content, lower oxygen consum ption rates and lower
free sulphide content to at least 2.5 cm depth. We
first tested if these differences, independent of
m echanical mixing by bioturbation, inhibited the
developm ent of the geochemical signature of elec­
trogenic sulphur oxidation. Sedim ent was collected
in May 2011, hom ogenized and defaunated by
passing it through a 500 pm sieve. The sedim ents
were then repacked into core tubes (4 cm diameter),
w ith the sedim ent surface adjusted to be level w ith
the top of the core liner. The cores were placed in an
aquarium w ith water draw n from the study site and
kept at 16 °C in a clim ate-controlled room. The
aquarium was continuously bubbled to ensure full
aeration, and water was replaced weekly. The
potential developm ent of electrogenic sulphur oxi­
dation in these sedim ents was tracked by recording
m icrosensor profiles in triplicate sedim ent cores
every few days.
The first experim ent confirmed that these sedi­
m ents could develop the geochemical signature of
electrogenic sulphur oxidation (see Results). Conse­
quently, we then designed a second m anipulation
experim ent to test if bioturbation specifically could
halt electrogenic sulphur oxidation. Intact sedim ent
cores were collected intact from OSF in August
2011. To accommodate bioturbating macrofauna,
these cores were 15 cm diam eter and had ~ 2 0 c m
sedim ent depth w ith 15 cm overlying water. Cores
were incubated in a climate controlled room at 16 °C
throughout the experim ent. Cores were first defau­
nated by asphyxia, w hich was achieved by closing
the cores w ith an air-tight sealing. After 10 days, the
cores were re-opened and dead m acrofauna (mostly
the lugworm A. marina) was carefully rem oved from
the sedim ent surface. These defaunated sedim ent
cores were then incubated w ith fully aerated water,
and we refer to this as Day 1 of the experiment. On
Day 8, m icrosensor profiles were recorded, and
revealed the geochemical signature of electrogenic
sulphur oxidation. On Day 10, the dom inant
bioturbating anim al (the lugworm A. marina) was
collected from the OSF. Two lugworms were
re-introduced to each sedim ent core, achieving
approximately natural densities (~ 1 0 0 m ~ 2). Extensive
m icrosensor profiling was conducted again on
Day 15 (referred to as ‘re-faunated’ cores). Vertical
m icrosensor profiles were m easured both in areas
where the sedim ent was not visibly disturbed, and
in areas where the sedim ent was heavily disturbed
by the activities of the lugworms (that is, around the
feeding pocket and the fecal casts). Extended
experimental details are available in Rao et al.
(in review).
Results
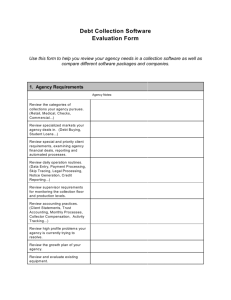
A geochemical signal of long-range electron transport
in natural sediments
The geochemical signature of electrogenic sulphide
oxidation was present at three of the four coastal zone
habitats investigated in the North Sea area. Represen­
tative microsensor profiles are shown for the three
sites where the geochemical signature was found
(RSM, BCZ and MLG; Figures la-c), and the one site
where it was not found (OSF; Figure Id; all site
locations are shown in Figures le and f). Furthermore,
the geochemical signature of electrogenic sulphur
oxidation was evident on multiple visits at the three
sites (RSM, BCZ and MLG), identifying that electro­
genic sulphide oxidation occurs regularly at these
sites. These three sites were characterized by high
DOU rates (range: 13.9-44.1 at RSM, BCZ and MLG vs
3.5-4.8mmol 0 2m ~ 2d ~ 1 at OSF; Table 1). The first
three sites had suboxic zones between 5 and 40 mm
wide and typically showed a steep increase in the
XH2S concentration below the suboxic zone. At these
three sites, at least 5.3-33.7% of the DOU was
attributable to electrogenic sulphide oxidation
(Table 1). Current densities ranged from 4.6 ±
1.8m A m ~ 2 at BCZ in March 2013 up to
29.9 ± 10.7m A m ~2 at RSM in June 2011.
Long Desulfobulbaceae filaments in natural sediments
At all sites w ith geochemical evidence of electro­
genic sulphide oxidation, m icroscopic exam ination
of the sedim ents revealed long, unbranched,
filam entous bacteria (Figure 2a), w hich were subse­
quently identified by CARD-FISH as Desulfobulbaceae
(Figure 2b; Supplem entary Figures S i and S2).
Enum eration using FISH revealed densities of 82.0
and 122.8 m of filament per cubic centim etre of
bulk sedim ent at RSM and BCZ, respectively.
No filaments were detected at the OSF reference
site, w here there was no geochemical evidence of
The ISME Journal
Long-range electron transport in the seafloor
SY M alkin et al
o
5
E 1°
E
.c 15
Q_
(D
°
20
25
30 -
r
T 1-- 1-- 1-- 1---1
100
200
300
0 2 or £H2S Omoi L 1)
The Netherlands
ÍS.
MLG
BCZ
Belgium
Figure 1 M icrosensor depth profiles of 0 2 (red), SH2S (blue) and pH (black) show the characteristic biogeochem ical fingerprint of
electrogenic su lp h u r oxidation in surface sedim ents from three coastal sites in the North Sea: (a) RSM, (b) BCZ and (c) MLG.
In contrast, m icrosensor depth profiles from (d) OSF do not show the characteristic fingerprint. R epresentative m icroprofile curves are
illustrated, (e) World map indicating A arhus Harbour, the location w here the electrogenic signature and Desulfobulbaceae filam ents
were first found in laboratory incubations (yellow cross), and the present study region (Dutch Delta area; red triangle), (f) Detailed map
of individual study site locations (red triangles); full site details are given in Table 1. The w orld map also indicates locations w here the
geochem ical signature of long distance electron transport have been found in literature reports (black circles) and locations w here
highly sim ilar (^ 9 7 % ) 16S rRNA gene sequences to the Desulfobulbaceae filam ents w ere retrieved from GenBank (blue squares); see
D iscussion for details.
electrogenic sulphide oxidation. Filam ent fragments
up to 8 mm in length (~ 4000 cells) were retrieved
from MLG sedim ents by gently dissecting sedim ent
under a microscope. SEM images revealed a pattern
of evenly spaced external ridges running in parallel
along the length of the filam ents isolated from all
three sites (Figure 2c; S upplem entary Figures S i
and S2). Cell diam eters were 1.25-3.04 pm
at RSM, 0 .8 9 -1 .3 7 pm in the BCZ and 2.182.36 pm in MLG based on m easurem ents of 2-5
filam ents m easured using SEM per site. Filam ents
w ith larger cell diam eters had more ridges and we
estim ated across all sam ples betw een 16 and 58
ridges per filam ent.
Desulfobulbaceae clones w ith ^ 9 7 % sequence
sim ilarity to the 16S rRNA gene sequences reported
from the laboratory incubations of Pfeffer et al.
(2012), were found in all field samples that showed
the electrogenic sulphur oxidation signature (RSM,
BCZ and MLG; Figure 2d). A summ ary of the full
clone library com position is given in Supplem entary
Figure 4. Sequences have been deposited in
GenBank
under
accession
num bers
BankI t l 707352:KJ562723-KJ562819. A clone library was
The ISME Journal
not constructed from OSF sedim ents, where the
geochemical signature of electrogenic sulphur oxi­
dation was not observed. However, a previous
study of sim ilar bioturbated sedim ents in
the Oosterschelde found no Desulfobulbaceae
sequences in either the surface-oxidized sedim ent
or the deeper sulphate-reducing sedim ent (Miyatake
et al., 2013).
Effects of bioturbation on geochemical signal
The geochemical fingerprint of electrogenic sulphur
oxidation was not present in intact sedim ent cores
collected from the highly bioturbated sandy sedi­
m ent at OSF collected in May and August 2011.
However, sedim ents from OSF that were defaunated
by sieving and incubated under darkened laboratory
conditions in aerated seawater developed the pH
profile indicative of electrogenic sulphur oxidation
(Figure 3). The characteristic pH profile indicative
of cathodic oxygen consum ption was first evident
on Day 5 after hom ogenization, whereby porew ater
pH was just beginning to increase at the base of the
oxic zone (Figure 3). This pH peak progressively
Long-range electron transport in the seafloor
et al
SY M alkin
^TRSM -B-20^
59 i-^-n-RSM-B-27
—I
RSM-B-22
*----------RSM-B-4
Aarhus (JX091073)
88
Aarhus (JX091072)
“ BCZ-B-15
J-MLG-B-27
vI r SM-B-6
0.05
“
Desulfobulbus japonicus
*
Desulfobulbus propionicus
—
D esulfovibrio alkaliphilus
Desulfocapsa sulfexigens
81
--------------------------
-C
"
Desulfobacter postgatei
' Desulfonema magnum
Desulfovibrio desulfuricans
Figure 2 Identification of filam entous Desulfobulbaceae bacteria present in intact sedim ent from sites described in Figure 1 and Table 1.
(a) A bundant long filam entous bacteria taken from near the surface of sedim ent and gently teased apart w ith forceps, (b) Identification of
the filam ents belonging to Desulfobulbaceae using CARD-FISH (DSB 706 probe), (c) SEM image of a bacterial filam ent isolated from the
sedim ent. All images show n here are from MLG. See Supplem entary m aterial for images from RSM and BCZ. (d) Phylogenetic tree of
Desulfobulbaceae 16S rRNA sequences recovered from intact sedim ent described in Figure 1. Closely related sequences of filam entous
Desulfobulbaceae from A arhus Bay and other isolated bacteria are also included. Scale bar shows 5% sequence divergence and bootstrap
support is indicated on tree branches.
increased over the days of incubation, w ith a
m axim um subsurface pH observed on Day 20.
At the start of the incubation, before the onset of
cathodic oxygen consum ption, the DOU declined
from 6.3 ±0.7 at Day 1 to 3.7 ± 0.1 mmol 0 2m -2 d -1
at Day 5. After Day 5, in concert w ith the develop­
m ent of a pH m axim um in the oxic zone, there was
an increase in DOU. Between Day 5 and Day 20,
DOU increased from 3.7 ±0.1 to 5.4 ±0.2 mmol
0 2m _2d _1, representing a 51% increase in oxygen
uptake. Over this same period (Days 5-20), the
estim ated electrical current density increased from
0.5 ±0.3 up to 9.5 ± 1 .4 m A m " 2. The percentage of
sedim ent DOU attributed to cathodic reactions
increased from 0% on Day 1 to 22.4 ± 1.2% on Day
5, and up to 51.6 ±3.7% on Day 20. Although
CARD-FISH was not performed during this
particular experiment, subsequent analysis of
sedim ent collected from OSF and sieved and
incubated
following
the
same
procedures,
The ISME Journal
Long-range electron transport in the seafloor
SY M alkin et al
a
pH
6.0
7.5
9.0 6.0
I .1. I .1 1
°5 - F
I
I
I
7.5
I
I
I
9.0 6.0
I
I
I
I
7.5
I
I
I
9.0 6.0
I
I
I
I
7.5
I
I
I
9.0 6.0
I
I
I
I
7.5
I
I
I
9.0 6.0
I
I
I
I
7.5
I
I
I
9.0
I
I
I
F
E 10-
E
£
15 H
25Day 1
Day 5
Day
30-
Day 13
1 r i i i i i i
0
100
250
0
100
250
0
100
Day 20
Day 47
i i i i i i i
250
0
100
250
0
100
d
60 -i
250
0
100
20
30
250
I H 2S or 0 2 (pm ol iL- 1 )\
7
12 -i
10
T3
'e
■a:
o
E 5
^E
3
o
Q
50-
-
6
E 8
<
E 6
40 -
^30Q.
4
20-
2
10-
4
3
1
10
20
30
40
Time from S tart (d)
50
0
10
20
30
40
50
Time from S tart (d)
0
10
40
50
Time from S ta rt (d)
Figure 3 (a) A subset of m icrosensor profiles from sedim ent collected from a heavily bioturbated sedim ent from an OSF that has been
artificially defaunated and incubated w ith well-oxygenated overlying water. Despite a low concentration of free sulphide, developm ent
of a geochemical signature of long-range electron transport is evident from Day 5 onwards, (b) DOU during the incubations, average of
n = 3, plotted w ith standard error about the mean, (c) Current density calculated from these sedim ents and (d) percentage, p, of oxygen
flux that is attributable to cathodic oxygen uptake.
confirmed the presence of Desulfobulbaceae
filaments w henever the geochemical signature of
electrogenic sulphur oxidation was present
(Supplem entary Figure 3).
To test if infauna specifically can inhibit the
geochemical signature of electrogenic sulphide
oxidation, we performed a second m anipulation
experiment, w here fauna were first removed by
asphyxiation and subsequently reintroduced. W hen
intact sedim ent cores were retrieved from the OSF
field site and im m ediately profiled, these sedim ents
did not exhibit evidence of electrogenic sulphur
oxidation (Figure 4a). In defaunated sedim ents, the
geochemical signature of electrogenic sulphur
oxidation again developed w ithin 8 days of exposure
to aerated seawater (Figure 4b). The deep burrowing
lugworm A. marina was then re-introduced and
caused extensive sedim ent overturning, removing
sedim ent from the feeding pocket and expelling
sedim ent and organic waste as fecal casts on the
sedim ent surface. In sedim ent patches that were not
m echanically disrupted by the burrow ing lugworms,
the electrogenic signature persisted, though w ith
attenuated pH extremes (Figure 4c). In sedim ent
patches that were directly overturned by lugworm
bioturbation, the electrogenic signature disappeared
entirely (Figure 4d).
The ISME Journal
Discussion
Our results provide evidence that electrogenic
sulphur oxidation, previously reported from labora­
tory incubations (Nielsen et al., 2010; Pfeffer et al.,
2012; Risgaard-Petersen et al., 2012), occurs under
natural conditions in the seafloor. We observed
evidence of electrogenic sulphur oxidation, together
w ith Desulfobulbaceae filaments, in a variety of
coastal m arine sedim ents (that is, a salt m arsh
drainage channel, a subtidal site recovering from
seasonal hypoxia, a coastal area of high m ud
deposition). These sedim ents were characterized
by high DOU and high organic m atter content.
In the narrow oxic zone (0.7-2.0 mm depth), a pH
m axim um was observed under dark conditions,
indicative of cathodic oxygen consum ption. Below
this depth, there was a suboxic zone where oxygen
and sulphide were not detectable, w hich was
typically 5-15 mm deep (although sometimes
extending deeper). Below the suboxic zone, these
sedim ents exhibited a steep sulphide gradient,
indicating high sulphide production. Repeated
visits to these three study sites identified that
electrogenic sulphur oxidation is a regularly occur­
ring, although not necessarily perm anent, feature at
these field sites.
Long-range electron transport in the seafloor
et al
SY M alkin
asphyxiation
fauna re-introduction
re-aeration
I
u
Cl) c(/)
c5
Cl) .Ç
£
T<D
1)
intact
defaunated
lightly disturbed
heavily disturbed
CO
d
pH
9
pH
6
ts
300
0 2 or IH 2S (^M)
0 2 or IH 2S (^M)
300 0
0 2 or SH2S (hM)
100
200
300
0 2 or IH 2S (hM)
Figure 4 Effect of sedim ent disturbance by fauna on electrogenic su lp h u r oxidation. Microprofiles of 0 2 (red), £H 2S (blue) and pH
(black and grey) are show n from sedim ent cores collected from the intertidal OSF. Different shades are used for replicate profiles for pH
and £H 2S for image clarity, whereas a single m edian profile is show n for 0 2. (a) Profiles from intact sedim ent cores, (b) Profiles from
defaunated sedim ent cores, (c) Profiles from re-faunated sedim ent cores, in areas that were not visibly disrupted by m echanical
disturbance caused by the re-introduced lugworms. (d) Profiles from re-faunated sedim ent cores, in areas that were not visibly disrupted
by m echanical disturbance caused by the re-introduced lugworms.
Our observation of the geochemical signature of
electrogenic sulphur oxidation always coincided
w ith the presence of Desulfobulbaceae filaments
that were closely related (that is, ^ 9 7 % 16S rRNA
gene sequence similarity) to those previously
reported from the laboratory incubations of Pfeffer
et al. (2012). Desulfobulbaceae filaments from the
suboxic zone of our study sites revealed densities of
82.0 and 122.8m filam ent per cubic centimetre
sedim ent at RSM and BCZ, respectively, and no
filaments were found at the reference site OSF.
These densities are sim ilar to those reported in
Pfeffer et al. (2012) (127 m c m -3) and support the
hypothesis that the long-range electron transport
associated w ith electrogenic sulphur oxidation is
m ediated by the Desulfobulbaceae filaments (Pfeffer
et al., 2012).
The filam entous bacteria retrieved from the three
electrogenic sulphur oxidation study sites (RSM,
BCZ and MLG) were long and unbranched, and
retrieved fragments could exceed eight m illim etres
in length. The unusual m orphological feature of
raised ridges running in parallel along the length
of the filaments, observed previously in laboratory
incubations,
were
also
prom inent in the
Desulfobulbaceae filaments isolated in this study.
Pfeffer et al. (2012) reported filament diameters
betw een 0.4 and 0.7 pm w ith fewer than 20 ridges
per filament, whereas we herein report filam ent
diameters betw een 0.9 and 3.0 pm w ith 16-58 ridges
per filament. It rem ains to be seen if these
morphological differences represent genotypic or
phenotypic variation.
A cosm opolitan distribution of electrogenic
sulphur oxidation is supported by a num ber of
published datasets in w hich the geochemical finger­
print of this process is evident, but has not been
formally recognized (Figure Id). Shadow subsurface
pH maxima, characteristic of cathodic oxygen con­
sum ption, have been reported from in situ m icro­
sensor profiling in the Santa Barbara (Cai et al.,
2000) and Santa M onica Basins (Reimers et al.,
1996). These basins are perm anently hypoxic,
support high rates of sulphate reduction, and are
devoid of bioturbating fauna. Similar pH profiles
were also reported from a site situated w ithin an
oyster aquaculture park in the Thau lagoon (Medi­
terranean Sea, France; Dedieu, 2005) and from
sedim ents near a fish farm in Pillan fjord (Chile;
M ulsow et al., 2006), both of w hich experience high
The ISME Journal
Long-range electron transport In th e seafloor
SY M alkin e f a/
organic m atter loading and having little or no
burrow ing infauna. Seasonally hypoxic sedim ents
from Tokyo Bay have also dem onstrated m icropro­
file evidence of cathodic oxygen consum ption
(Sayama, 2011), sim ilar to those reported here from
MLG. At a site characterized by high sulphide fluxes
and devoid of large bioturbating infauna on the MidA tlantic ridge, high rates of sulphide oxidation were
detected w hich could not be attributed to any other
know n sulphide removal process, such as sulphur
oxidation by large nitrate-accum ulating bacteria
(Schauer et al., 2011). One explanation is that
electrogenic sulphur oxidations is responsible for
the observed sulphide removal, however, direct
evidence is needed to confirm this possibility.
Gene sequence archives also support a cosm opo­
litan distribution of the conductive filamentous
bacteria in locations w ith high rates of sulphide
generation, via high organic m atter loading or
sulphide seepage, and a paucity of bioturbating
animals. 16S rRNA gene sequences that were highly
sim ilar
97%) to the Desulfobulbaceae at our field
sites have been detected at cold seeps on the
Hikurangi Margin (Baco et al., 2010) and the Nile
Deep Sea Fan (Griinke et al., 2011) in areas devoid of
fauna; in organic rich cohesive sedim ent of a
subtropical mangrove swam p (Liang et al., 2007);
and at a sewage im pacted site in the Bay of Cadiz
(Köchling et al., 2011).
Previous laboratory experim ents have show n that
sulphur oxidation by long-range electron transport
can play a dom inant role in oxygen consum ption
and can exert a strong im print on the overall sulphur
cycling and pH dynamics in sedim ents (Nielsen
et al., 2010; Risgaard-Petersen et al., 2012). Our
observations show that this process likely has a
sim ilarly profound effect on m ineral cycling in some
natural coastal sedim ents. We observed pH values as
high as 8.8 in the oxic zone, and as low as 6.1 in the
suboxic zone (Figure 1). These pH extremes lead
to dissolution of metal sulphides and calcium
carbonate w ithin the deep suboxic zone, followed
by re-precipitation of iron (hydr)oxides and
carbonate near the sedim ent-w ater interface
(Risgaard-Petersen et al., 2012). High alkalinity
effluxes have been reported from m uddy sites on
the BCZ, including Station 130 (Braeckman et al.,
2014), and such effluxes are consistent w ith
cathodic oxygen consum ption and elevated carbo­
nate dissolution induced by electrogenic sulphur
oxidation. Current densities estim ated at the three
field sites dem onstrating electrogenic sulphur
oxidation (15-23 m A m -2) are comparable w ith the
values observed in previous laboratory experim ents
(Nielsen et al., 2010; Risgaard-Petersen et al., 2012)
and are sim ilar to those reported from the anodes of
sedim ent batteries colonized by natural microbial
com m unities (Tender et al., 2002; Ryckelynck et al.,
2006). We estim ated that a m inim um of 5.3-33.7%
of the DOU in these sedim ents was due to electro­
genic sulphur oxidation, substantiating that this
The ISME Journal
process can be a dom inant biogeochemical process
in natural m arine sedim ents.
Our data further suggest that bioturbation
may exert a major control on the natural distribution
of electrogenic sulphur oxidation in coastal m arine
sediments. In heavily bioturbated sediment from OSF,
the geochemical signature of electrogenic sulphide
oxidation was not detected under field conditions, but
could be induced in laboratory experiments, as long as
bioturbation was excluded. In these sediments, a seed
population of Desulfobulbaceae bacteria m ust have
existed, from w hich these filam ents w ere able to
proliferate w hen burrow ing anim als are rem oved.
We observed that sedim ent overturning by
bioturbation caused a clear disru p tio n of the
electrochem ical signal. This m ay arise from
a direct breakage of the bacterial filam ents, in a
m anner sim ilar to the previously described cutting
experim ents of Pfeffer et al. (2012). In addition,
sedim ent overturning m ay halt electrogenic
su lp h u r oxidation by depositing an anoxic layer
of sedim ent on top of the sedim ent surface, thereby
depriving the resident filam entous bacteria from
access to electron acceptors.
In summary, we have shown that electrogenic
sulphur oxidation is found in intact coastal marine
sedim ents under natural conditions. We found this
process, together w ith the Desulfobulbaceae fila­
ments, in three depositional sedim ent areas (a salt
marsh, a subtidal coastal m ud plain and a hypoxic
marine basin). These sediments were collectively
characterized as being rich in organic matter, and
consequently supporting high rates of sulphate
reduction, and having an oxygenated overlying water
column. These sedim ents were furthermore subject
to little biomechanical mixing by macrofauna, which
appears to be a key control on the distribution of
electrogenic sulphur oxidation in natural settings.
Until now, the occurrence of electrogenic sulphur
oxidation has not been accounted for in studies of
natural marine sediments. Given its w idespread
distribution, electrogenic sulphur oxidation may be
a key process in the biogeochemistry and microbial
ecology of the seafloor.
Conflict of Interest
The authors declare no conflict of interest.
Acknowledgements
We th a n k U B raeckm an for p ro v id in g d ata on the
m acro fau n a d istrib u tio n in BCZ, R D asseville (U Gent)
for h elp w ith SEM, V C onfurius for assistan ce w ith
clone lib raries an d S E ng elh ard (NIOZ) for h elp
w ith CA RD-FISH im ages. We th a n k th ree an onym ous
review ers for th e ir com m ents. T his research w as
fin an cially su p p o rte d by th e B russels In stitu te for
R esearch an d In n o v atio n (H oloFlow project: SYM, EMZ),
th e D arw in C enter for B iogeosciences (DS, DV-C, HTSB),
th e R esearch F o u n d atio n - F lan d ers (FW O -O dysseus grant
Long-range electron transport In the seafloor
SY M alkin e ta /
FJRM), th e N eth erlan d s O rganisation for S cientific
R esearch (NWO-VIDI grant FJRM) an d the E uro p ean
R esearch C ouncil (ERC S tarting grant FJRM).
References
A sh elford KE, C h u zhanova NA, Fry JC, Jones AJ,
W eightm an AJ. (2005). At least 1 in 20 16S rRNA
sequence records cu rren tly h e ld in p u b lic reposito ries
is estim ated to co n tain su b stan tia l anom alies. A p p l
E nviron M icrobiol 71: 7724-7736.
Baco AR, R ow den AA, L evin L, S m ith CR, B ow den DA.
(2010). Initial ch aracterizatio n of cold seep faunal
com m u n ities on the N ew Z ealan d M argin. M ar Geol
272: 251-259.
B o u d reau BR (1996). The diffusive to rtu o sity of fin e­
g rain ed u n lith ifie d sedim ents. G eochim C osm ochim
A d a 60: 3139-3142.
B raeckm an U, Foshtom i MY, Van G ansbeke D, M eysm an F,
Soetaert K, V incx M et al. (2014). Variable im portance
of m acrofaunal functional biodiversity for biogeochem ­
ical cycling in tem perate coastal sedim ent. E cosystem s
(in press); doi:10.1007/sl0021-014-9755-7.
Clarke TA, E dw ards MJ, G ates AJ, H aii A, W hite GF,
B radley J et al. (2011). S tru ctu re of a bacterial cell
surface d ecahem e electron con d u it. Proc N a t A c a d S ei
108: 9384-9389.
Cai W-J, Z hao R W ang Y. (2000). pH a n d p C 0 2 m icro elec­
trode m e asu rem en ts an d th e diffusive beh av io r of
carbon dio x id e sp ecies in coastal m arin e sedim en ts.
M ar C hem 70: 133-148.
C oosen J, Seys J, M eire PM, C raeym eersch J. (1994).
Effect of sedim entological an d h y d ro d y n am ical
changes in th e in te rtid a l areas of th e O ostersch eld e
estuary (SW N etherlands) on d istrib u tio n , d en sity
an d
biom ass
of five
com m on
m acroben th ic
species: S pio m artin en sis (M esnil), H ydrobia ulvae
(Pennant), A renicola m arin a (L.), S coloplos arm iger
(M uller) an d B athyporeia sp. H ydrobiologia 282/283:
235-249.
D edieu K. (2005). B iogeochem ical dy n am ics of oxygen in
coastal sedim ents: stu d y by m icro-electrodes an d
m odels. Ph.D. thesis. U niversité Pierre et M arie
C urie (Paris), France. N ational th esis num ber:
2005PA066019, 2005.
D ickson AG, Sabine CL, C hristian JR (eds.) (2007). G uide
to B est P ra d ic es For O cean C 0 2 M easurem ents. PICES
S pecial P u b licatio n 3, p p 191.
Gao Y, Lesven L, G illan D, Sabbe K, B illon G, De C alan S
et al. (2009). G eochem ical b ehavior of trace elem ents
in sub-tidal m arin e se d im en ts of th e Belgian coast.
M ar C hem 117: 88-96.
C orby YA, Y anina S, M cLean JS, Rosso KM, M oyles D,
D ohnalkova A et al. (2006). E lectrically co ndu ctiv e
b acterial n an o w ires
p ro d u c e d
by
S hew an ella
o n eid en sis strain MR-1 a n d other m icroorganism s.
Proc N a tl A c a d S d 103: 11358-11363.
G riinke S, F eld e n J, L ichtschlag A, G irnth A-C, De Beer D,
W enzhöfer F et al. (2011). N iche differen tiatio n am ong
m at-form ing, sulfide-oxidizing bacteria at cold seeps
of th e N ile D eep Sea F an (Eastern M e d iterran ean Sea).
G eobiol 9: 300-348.
H ofm ann AF, S oetaert K, M id delburg JJ, M eysm an FJR.
(2010). A quaEnv: an aquatic a c id -b a se m odellin g
e n v iro n m en t in R. A q u a t G eochem 16: 507-546.
Jorgensen BB. (1982). M in eralizatio n of organic m atter in
th e sea b ed - th e role of su lp h a te red u ctio n . N ature
296: 643-645.
Jorgensen BB, N elso n DC. (2004). S ulfide o x id atio n in
m arin e sed im en ts: g eochem istry m eets m icrobiology.
In: A m en d JP, E d w ard s K, Lyons TW (eds). S u lfu r
B iogeochem istry—P ast a n d Present, vol 379. Special
Paper, G eological S ociety of A m erica, p p 63-81.
K öchling T, L ara-M artin P, G onzalez-M azo E, A m ils R,
Sanz JL. (2011). M icrobial co m m u n ity co m p o sitio n of
anoxic m arin e se d im en ts in th e Bay of C adiz (Spain).
In tern a t M icrobiol 14: 143-154.
Liang J-B, Chen Y-Q, Lan C-Y, Tam NF Y, Z an Q-J, H uang L-N.
(2007). R ecovery of no v el b acterial d iv ersity from
m angrove sed im en t. M ar B iol 150: 739-747.
Logan BE, Rabaey K. (2012). C onversion of w astes in
b io electricity an d chem icals u sin g m icro b ial electro ­
ch em ical technologies. S d e n c e 337: 686-690.
Lovley DR. (2008). T he m icrobe electric: co n v ersio n of
organic m atter to electricity. Curr O pin B iotech 19:
564-571.
Liicker S, Steger D, Kjeldsen KU, MacGregor BJ, Wagner M,
Loy A. (2007). Im p ro v ed 16S rR N A -targeted probe
set for analysis of su lfate-red u cin g b acteria by flu o r­
escence in situ h y b rid izatio n . J M icrobiol M eth o d s 69:
523-528.
L udw ig W, S tru n k O, W estram R, R ichter L, M eier H,
Y adhukum ar et al. (2004). ARB: a software environm ent
for sequence data. N u d e ic A d d s Bes 32: 1362-1371.
M iyatake T, M acG regor BJ, B oschker HTS. (2013).
D ep th -related differences in organic su b strate u tiliz a ­
tio n by m ajor m icro b ial g roups in in te rtid a l m arin e
sed im en t. A p p l E nviron M icrobiol 79: 389-392.
M ulsow S, K rieger Y, K en n ed y R. (2006). S ed im en t profile
im aging (SPI) an d m icro -electro d e tech n o lo g ies in
im p act assessm en t stu d ies: ex am ple from tw o fjords in
S o u th ern C hile u se d for fish farm ing. J M ar S y st 62:
152-163.
N ielsen LP, Risgaard-Petersen N, Fossing H, C hristensen PB,
Sayam a M. (2010). E lectric cu rren ts co u p le sp atially
sep arated biogeochem ical p ro cesses in m arin e
sed im en t. N ature 463: 1071-1074.
O enem a O. (1988). Early diagenesis in recen t fine-grained
se d im en ts in th e E astern S ch eld t, Ph.D. thesis, U trecht
U niversity: U trecht, T he N eth erlan d s.
P a llu d C, Van C ap p ellen P. (2006). K inetics of m icrobial
sulfate re d u c tio n in estu arin e sed im en ts. G eochim
C osm ochim A d a 70: 1148-1162.
P ern th aler A, P ern th aler J, A m an n R. (2002). F lu o rescen ce
in situ h y b rid iz atio n an d cataly zed rep o rted
d ep o sitio n for th e id e n tifica tio n of m arin e bacteria.
A p p E nviron M icrobiol 68: 3094-3101.
Pfeffer C, Larsen S, Song J, Dong M, Besenbacher F, M eyer RL
et al. (2012). F ilam en to u s b acteria tran sp o rt electrons
over cen tim eter d istan ces. N ature 491: 218-221.
Rao AMF, M alkin SY, M eysm an FJR. (in review ).
E n h an c ed alk alin ity p ro d u c tio n in in te rtid a l san d s
from th e O o stersch eld e (The N eth erlan d s) in d u c e d by
th e lugw orm A ren ico la m arina. E stuarine Coast S h e lf
S ei (ECSS-S-13-00698; p p 46).
Reguera G, M cCarthy KD, M ehta T, Nicoli JS, Tuom inen MT,
Lovley DR. (2005). E x tracellu lar electro n tran sfer via
m icro b ial n an o w ires. N a tu re 435: 1098-1101.
R eguera G, N evin KP, N icoli JS, C ovalla SF, W oodard TL,
Lovley DR. (2006). B iofilm an d n an o w ire p ro d u ctio n
leads to increased current in Geobacter su lphurreducens
fuel cells. A p p l Environ Microbiol 72: 7345-7348.
The ISME Journal
Long-range electron transport In th e seafloor
SY M alkin e f a/
Reimers CE, Ruttenberg KC, Canfield DE, C hristiansen MB,
M artin JB. (1996). Porew ater pH and authigenic phases
form ed in the upperm ost sedim ents of the Santa Barbara
Basin. G eochem Cosmo A cta 60: 4 0 3 7 ^ 0 5 7 .
R isgaard-Petersen N, Revil A, M eister P, N ielsen LP.
(2012). S u lp h u r, iro n a n d calcium cycling asso ciated
w ith n atu ra l electric cu rren ts ru n n in g th ro u g h m arine
sedim ents. G eochim C osm ochim A d a 92: 1-13.
Ryckelynck N, S techer III HA, R eim ers CE. (2006).
U n d ersta n d in g the ano d ic m e ch an ism of a seafloor
fuel cell: in te ractio n s b etw een geochem istry an d
m icrobial activity. B iogeochem 76: 113-139.
S avitzky A, G olay MJE. (1964). S m oothing an d d ifferen tia­
tio n of d ata by sim p lified least squares pro ced u res.
A n a l C hem 36: 1627-1639.
Sayam a M. (2011). Seasonal d ynam ics of su lp h id e
o x id atio n p rocesses in Tokyo Bay d ead zone se d i­
m ents. G old sch m id t C onference A bstracts.
S ch au er R, Roy H, A u g u stin N, G en n erich H-H, P eters M,
W enzhoefer F et al. (2011). B acterial s u lp h u r cycling
sh ap es m icro b ial co m m u n ities in surface se d im en ts of
an ultram afic h ydrotherm al vent field. Environ M icrobiol
13: 2633-2648.
S oetaert K, P etzo ld t T, M eysm an FJR. (2012). Package
‘m arelac’: Tools for aq u atic sciences, v.2.1.2.
T ender LM, R eim ers CE, S tech er III HA, H olm es DE, Bond
DR, Low y DA et al. (2002). H arn essin g m icrobially
g en erated p o w er on th e seafloor. N a t B io tech n o l 20:
821-825.
@ ® @ © This work is licensed under a Creative
L
J Commons Attribution-NonCommercialNoDerivs 3.0 Unported License. To view a copy
of this license, visit http://creativecommons.org/
licenses/by-nc-nd/3.0/
Supplementary Information accompanies this paper on The ISME Journal website (http://www.nature.com/ismej)
The ISME Journal